Дифференцировка лимфоцитов
2. Доантигенная дифференцировка лимфоцитов
Родоначальной клеткой Т-лимфоцитов, как и всех клеток крови, является полипотентная стволовая гемопоэтическая клетка. Её маркером является CD 34. Ранние предшественники Т-лимфоцитов мигрируют из костного мозга в тимус, где происходит антигеннезависимая дифференцировка Т-клеток под влиянием «клеток нянек», эпителиальных клеток тимуса, а так же гормонов тимуса (α- и β-тимозины, тимулин /сывороточный фактор тимуса/, тимопоэтин, тимический гуморальный фактор). Самыми ранними маркерами тимоцитов являются CD7, CD2. В тимусе Т-лимфоциты дифференцируются в иммунокомпетентные клетки и приобретают важную способность к распознаванию антигена. На их наружной мембране появляется (экспрессируется) особый рецептор - Т- клеточный рецептор (ТКР, англ. - TcR, T-cell receptor) для антигена.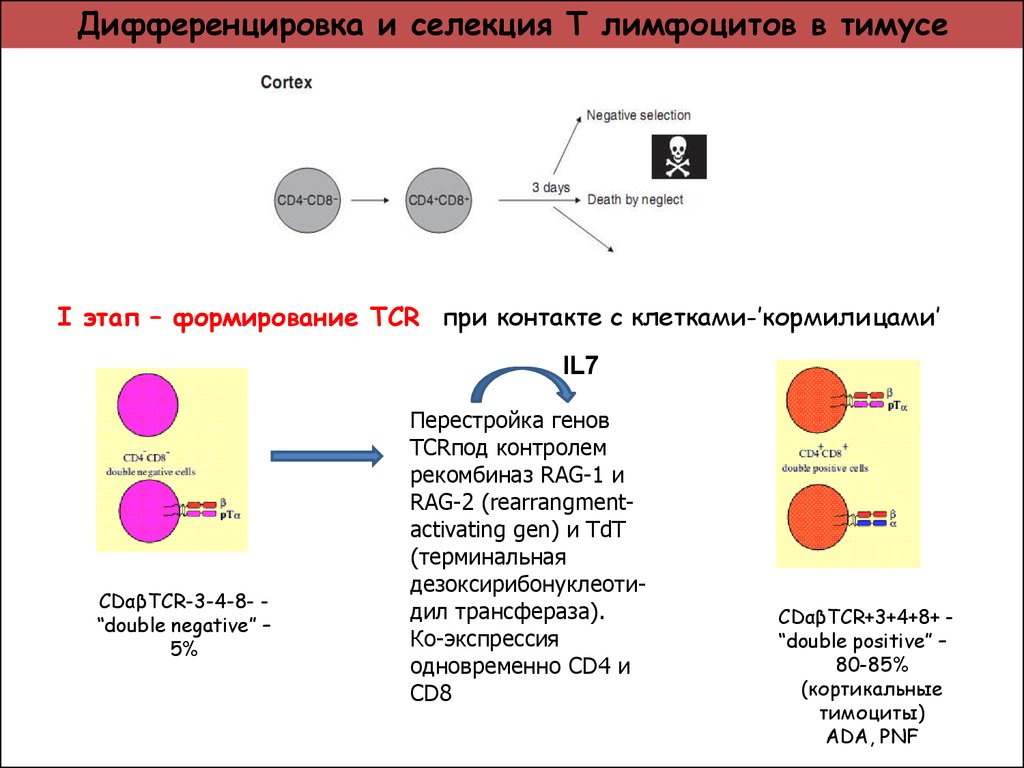
Причем для каждого антигена (эпитопа) в организме предназначен отдельный лимфоцит или его клональные дочерние лимфоциты-потомки, которые имеют специфичный антигену TcR. Тимоциты одновременно с TcR в процессе дифференцировки приобретают CD3, который тесно связан с Т-клеточным рецептором. CD3 необходим для передачи сигнала от ТКР в цитоплазму. На поверхности тимоцитов появляются также молекулы CD8 и CD4. Это двойные позитивные клетки, т.е. их фенотип (ТКР+, CD3+, CD4+, CD8+) и они являются наивными лимфоцитами.


В-лимфоциты также происходят от плюрипотентных гемопоэтических стволовых клеток, дающих также начало всем клеткам крови. Стволовые клетки находятся в определённом микроокружении, которое обеспечивает их выживание, самообновление или, при необходимости, дифференцировку. Микроокружение определяет, по какому пути пойдёт развитие стволовой клетки (эритроидному, миелоидному или лимфоидному).
Дифференцировка В-лимфоцитов, подобно Т-лимфоцитам, условно делится на две стадии — антигеннезависимую (в которую происходит перестройка генов иммуноглобулинов и их экспрессия) и антигензависимую (при которой происходит активация, пролиферация и дифференцировка в плазматические клетки). Выделяют следующие промежуточные формы созревающих В-лимфоцитов:
Ранние предшественники В-клеток — не синтезируют тяжёлых и лёгких цепей иммуноглобулинов, содержат зародышевые гены IgH и IgL, но содержат антигенный маркер, общий со зрелыми пре-В-клетками.
Ранние про-В-клетки — D-J-перестройки в генах IgН.
Поздние про-В-клетки — V-DJ-перестройки в генах IgН.
Большие пре-В-клетки — гены IgН VDJ-перестроены; в цитоплазме есть тяжёлые цепи класса μ, экспрессируется пре-В-клеточный рецептор.
Малые пре-В-клетки — V-J-перестройки в генах IgL; в цитоплазме есть тяжёлые цепи класса μ.
Малые незрелые В-клетки — гены IgL VJ-перестроены; синтезируют тяжёлые и лёгкие цепи; на мембране экспрессируются иммуноглобулины (B-клеточный рецептор).
Зрелые В-клетки — начало синтеза IgD.
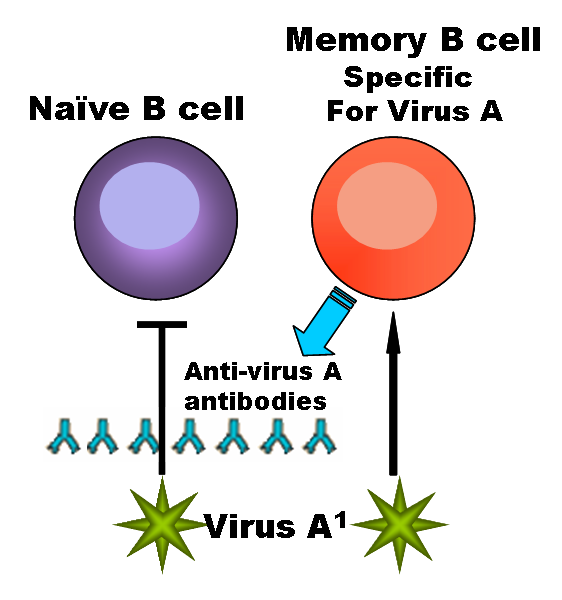
В-клетки поступают из костного мозга во вторичные лимфоидные органы (селезёнку и лимфатические узлы), где происходит их дальнейшее созревание, презентация антигена, пролиферация и дифференцировка в плазматические клетки и В-клетки памяти.
Экспрессия всеми В-клетками мембранных иммуноглобулинов позволяет осуществляться клональному отбору под действием антигена. При созревании, стимулировании антигеном и пролиферации существенно меняется набор маркеров В-клеток. По мере созревания В-клетки переключаются от синтеза IgM и IgD на синтез IgG, IgA, IgE (при этом у клеток сохраняется способность синтезировать также IgM и IgD — вплоть до трёх классов одновременно). При переключении синтеза изотипов антигенная специфичность антител сохраняется. Различают следующие типы зрелых В-лимфоцитов:
Собственно В-клетки (ещё называемые «наивными» В-лимфоцитами) — неактивированные В-лимфоциты, не контактировавшие с антигеном. Не содержат тельца Голла, в цитоплазме рассеяны монорибосомы. Полиспецифичны и имеют слабое сродство к многим антигенам.