Молекулярная биология регуляции артериального давления и фармакологические мишени управления гемодинамикой.
АПФ - типичная фармакологическая мишень для лечения ГБ
| Сайт: | Электронный информационно- образовательный портал ВолгГМУ |
| Курс: | Дисциплина Медицинская биохимия. Принципы измерительных технологий в биохимии. Патохимия, диагностика. Биохимия злокачественного роста». Часть 3 |
| Книга: | Молекулярная биология регуляции артериального давления и фармакологические мишени управления гемодинамикой. |
| Напечатано:: | Гость |
| Дата: | Вторник, 21 Май 2024, 06:50 |
Оглавление
- 1. Предисловие
- 2. Введение в регуляцию гемодинамики
- 3. Гуморальная регуляция тонуса гладких мышц сосудов и ОЦК
- 3.1. Ренин, ангиотензин, альдостероновая система (РААС или РАС)
- 3.2. Продукция и секреция ренина
- 3.3. Ангиотензиноген, ангиотензины 1,2,3,4 и т.д.
- 3.4. Реакции, катализируемые АПФ
- 3.5. Уровень АПФ в крови и особенности генетики человека
- 3.6. АПФ как мишень фармакологического воздействия
- 3.7. Классификация ингибиторов АПФ
- 3.8. Функциональная активность ангиотензина II
- 3.9. Ангиотензиновые рецепторы
- 3.10. Ангиотензиновые рецепторы как мишень фармакологического воздействия.
- 3.11. Неприлизин
- 4. Эндотелий-зависимые факторы
1. Предисловие
В регуляции артериального давления у человека участвует ренин-ангиотензиновая система (РАС). Работа РАС тесно связана с электролитами, они поддерживают гомеостаз, что необходимо для регуляции сердечной функции, баланса жидкости и многих других процессов. Один из компонентов РАС-системы – гормон ангиотензин II, который вызывает сужение сосудов, повышение артериального давления и является основным регулятором синтеза альдостерона, образующегося в клубочковой зоне коры надпочечников, единственного поступающего в кровь минералокортикоида человека. Конечным результатом такого действия является увеличение объема циркулирующей крови и повышение системного артериального давления.
2. Введение в регуляцию гемодинамики
Регуляция кровяного давления в норме
В соответствии с законами гидравлики АД прямо пропорционально произведению кровотока (сер дечный выброс, СВ) и сопротивления при прохождении крови через прекапиллярные артериолы (периферическое сосудистое сопротивление, или общее периферическое сопротивление — ОПС):
АД = СВхОПС.
Физиологически как у здорового, так и больного человека АД поддерживается путем регуляции CB и ОПС в трех анатомических областях:
- артериолах,
- посткапиллярных венулах (емкостные сосуды)
- сердце.

Рис. 1. Анатомические области, участвующие в регуляции кровяного давления
Четвертая анатомическая контрольная область — почки — вносит весомый вклад в поддержание АД, регулируя объем внутрисосудистой жидкости. Барорефлексы, реализуемые через симпатические нервы, совместно с гуморальными механизмами, включая ренин-ангиотензин-альдостероновую систему, координируют функции этих четырех контролирующих зон в под держании нормального АД. АД у больного с гипертензией регулируется теми же механизмами, что и у здорового человека с нормальными значениями АД. Регуляция АД при гипертензии отличается от нормы тем, что барорецепторы и почечный контроль давления, сопряженный с объемом крови, имеют “установочную точку” регуляции на более высоком уровне АД. Все антигипертензивные средства воздействуют на эти нормальные механизмы.
А. Постуральный барорефлекс (рис. 2). Барорефлексы участвуют в быстрой сиюминутной регуляции АД, например при переходе из горизонтального в вертикальное положение. Центральные симпатические нейроны, берущие начало в вазомоторной зоне продолговатого мозга, тонически активны. Каротидные барорецепторы стимулируются при растяжении стенки сосудов, вызванном давлением изнутри (АД). Активация барорецепторов тормозит центральные симпатические импульсы. Напротив, уменьшение растяжения ведет к снижению барорецепторной активности.

Рис. 2. Рефлекторная дуга барорефлекса
Таким образом, при переходе в вертикальное положение давление на барорецепторы снижается вследствие депонирования крови в венах ниже уровня сердца и симпатическая активность растормаживается. Рефлекторно возрастают периферическое сосудистое сопротивление (сокращение артериол) и сердечный выброс (прямая стимуляция сердца и сокращение емкостных сосудов, что ведет к увеличению венозного возврата к сердцу), и в результате восстанавливается нормальное АД. Такой же барорефлекс срабатывает в ответ на любое снижение АД, в том числе на первичное понижение периферического сосудистого сопротивления (например, вызванного сосудорасширяющим веществом) или уменьшение внутрисосудистого объема (например, из-за кровопотери или потери солей и воды через почки).
Б. Реакция почек на снижение АД. Почки участвуют в долгосрочной регуляции АД. Снижение почечной перфузии вызывает внутрипочечное перераспределение кровотока и повышает реабсорбцию соли и воды. Кроме того, снижение давления в почечных артериолах, также как и симпатическая активность (через b-адренорецепторы), стимулирует выработку ренина, который усиливает продукцию ангиотензина II. Ангиотензин II вызывает прямое сокращение резистивных сосудов и стимулирует синтез альдостерона в коре надпочечников, что ведет к повышению реабсорбции натрия в почках и повышению внутрисосудистого объема крови.2.1. Рефлекторная (быстрая) регуляция кровообращения
В первую очередь связана с постуральным барорефлексом, который перераспределяет кровоток при каждом изменении положения тела. Он реализуется в результате взаимодействия симпатической и парасимпатической нервных систем, степень активности которых регулирует сосудодвигательный центр, расположенный на дне 4 желудочка.
2.2. Вегетативная нервная система
Вегетати́вная не́рвная систе́ма (от лат. vegetatio — возбуждение, от лат. vegetativus — растительный), ВНС, автономная нервная система, ганглионарная нервная система (от лат. ganglion — нервный узел), висцеральная нервная система (от лат. viscera — внутренности), органная нервная система, чревная нервная система, systema nervosum autonomicum (PNA) — часть нервной системы организма, комплекс центральных и периферических клеточных структур, регулирующих функциональный уровень организма, необходимый для адекватной реакции всех его систем.
Вегетативная нервная система — отдел нервной системы, регулирующий деятельность внутренних органов, желез внутренней и внешней секреции, кровеносных и лимфатических сосудов. Играет ведущую роль в поддержании постоянства внутренней среды организма и в приспособительных реакциях всех позвоночных.
Анатомически и функционально вегетативная нервная система подразделяется на симпатическую, парасимпатическую и метасимпатическую. Симпатические и парасимпатические центры находятся под контролем коры больших полушарий, гипоталамических центров и центров продолговатого мозга.

Рис.3. Вегетативная нервная система.
В симпатическом и парасимпатическом отделах имеются центральная и периферическая части. Центральную часть образуют тела нейронов, лежащих в спинном и головном мозге. Эти скопления нервных клеток получили название вегетативных ядер. Отходящие от ядер волокна, вегетативные ганглии, лежащие за пределами центральной нервной системы, и нервные сплетения в стенках внутренних органов образуют периферическую часть вегетативной нервной системы.
Симпатические ядра расположены в спинном мозге. Отходящие от него нервные волокна заканчиваются за пределами спинного мозга в симпатических узлах, от которых берут начало нервные волокна. Эти волокна подходят ко всем органам.
Парасимпатические ядра лежат в среднем и продолговатом мозге и в крестцовой части спинного мозга. Нервные волокна от ядер продолговатого мозга входят в состав блуждающих нервов. От ядер крестцовой части нервные волокна идут к кишечнику, органам выделения.
Метасимпатическая нервная система представлена нервными сплетениями и мелкими ганглиями в стенках пищеварительного тракта, мочевого пузыря, сердца и некоторых других органов.
Деятельность вегетативной нервной системы не зависит от воли человека. Это означает, что в обычных условиях человек не может волевым усилием заставить сердце биться реже или мышцы желудка — не сокращаться. Однако достичь сознательного влияния на многие параметры, контролируемые ВНС, можно с помощью специальных методов тренировки — например, с использованием методов биологической обратной связи.
Симпатическая нервная система усиливает обмен веществ, повышает возбуждаемость большинства тканей, мобилизует силы организма на активную деятельность. Парасимпатическая система способствует восстановлению израсходованных запасов энергии, регулирует работу организма во время сна.
Под контролем автономной системы находятся органы кровообращения, дыхания, пищеварения, выделения, размножения, а также обмен веществ и рост. Фактически эфферентный отдел ВНС осуществляет нервную регуляцию функций всех органов и тканей, кроме скелетных мышц, которыми управляет соматическая нервная система.
В отличие от соматической нервной системы, двигательный эффекторный нейрон в автономной нервной системе находится на периферии, и спинной мозг лишь косвенно управляет его импульсами.
Морфология
Выделение автономной (вегетативной) нервной системы обусловлено некоторыми особенностями её строения. К этим особенностям относятся следующие:
- очаговость локализации вегетативных ядер в ЦНС;
- скопление тел эффекторных нейронов в виде узлов (ганглиев) в составе вегетативных сплетений;
- двухнейронность нервного пути от вегетативного ядра в ЦНС к иннервируемому органу.
Волокна автономной нервной системы выходят не сегментарно, как в соматической нервной системе, а из трёх отстоящих друг от друга ограниченных участков мозга: черепного, грудинопоясничного и крестцового.
Автономную нервную систему разделяют на симпатическую, парасимпатическую и метасимпатическую части. В симпатической части отростки спинномозговых нейронов короче, ганглионарные длиннее. В парасимпатической системе, наоборот, отростки спинномозговых клеток длиннее, ганглионарных короче. Симпатические волокна иннервируют все без исключения органы, в то время как область иннервации парасимпатических волокон более ограничена.
Автономная (вегетативная) нервная система подразделяется по топографическому признаку делятся на центральный и периферический отделы.
Центральный отдел
- парасимпатические ядра 3, 7, 9 и 10 пар черепных нервов, лежащие в мозговом стволе (краниобульбарный отдел), ядра, залегающие в сером веществе трёх крестцовых сегментов (сакральный отдел);
- симпатические ядра, расположенные в боковых рогах тораколюмбального отдела спинного мозга.
Периферический отдел
- вегетативные (автономные) нервы, ветви и нервные волокна, выходящие из головного и спинного мозга;
- вегетативные (автономные, висцеральные) сплетения;
- узлы (ганглии) вегетативных (автономных, висцеральных) сплетений;
- симпатический ствол (правый и левый) с его узлами (ганглиями), межузловыми и соединительными ветвями и симпатическими нервами;
- концевые узлы (ганглии) парасимпатической части вегетативной нервной системы.
Симпатический, парасимпатический и метасимпатический отделы
На основании топографии вегетативных ядер и узлов, различий в длине аксонов первого и второго нейронов эфферентного пути, а также особенностей функции вегетативная нервная система подразделяется на симпатическую, парасимпатическую и метасимпатическую.
Расположение ганглиев и строение проводящих путей
Нейроны ядер центрального отдела вегетативной нервной системы — первые эфферентные нейроны на пути от ЦНС (спинной и головной мозг) к иннервируемому органу. Нервные волокна, образованные отростками этих нейронов, носят название предузловых (преганглионарных) волокон, так как они идут до узлов периферической части вегетативной нервной системы и заканчиваются синапсами на клетках этих узлов. Преганглионарные волокна имеют миелиновую оболочку, благодаря чему отличаются беловатым цветом. Они выходят из мозга в составе корешков соответствующих черепных нервов и передних корешков спинномозговых нервов.
Вегетативные узлы (ганглии): входят в состав симпатических стволов (есть у большинства позвоночных, кроме круглоротых и хрящевых рыб), крупных вегетативных сплетений брюшной полости и таза, располагаются в области головы и в толще или возле органов пищеварительной и дыхательной систем, а также мочеполового аппарата, которые иннервируются вегетативной нервной системой. Узлы периферической части вегетативной нервной системы содержат тела вторых (эффекторных) нейронов, лежащих на пути к иннервируемым органам. Отростки этих вторых нейронов эфферентного пути, несущих нервный импульс из вегетативных узлов к рабочим органам (гладкая мускулатура, железы, ткани), являются послеузелковыми (постганглионарными) нервными волокнами. Из-за отсутствия миелиновой оболочки они имеют серый цвет. Постганглионарные волокна автономной нервной системы в большинстве своем тонкие (чаще всего их диаметр не превышает 7 мкм) и не имеют миелиновой оболочки. Поэтому возбуждение по ним распространяется медленно, а нервы автономной нервной системы характеризуются бо́льшим рефрактерным периодом и большей хронаксией.
Рефлекторная дуга
Строение рефлекторных дуг вегетативного отдела отличается от строения рефлекторных дуг соматической части нервной системы. В рефлекторной дуге вегетативной части нервной системы эфферентное звено состоит не из одного нейрона, а из двух, один из которых находится вне ЦНС. В целом простая вегетативная рефлекторная дуга представлена тремя нейронами.
Первое звено рефлекторной дуги — это чувствительный нейрон, тело которого располагается в спинномозговых узлах и в чувствительных узлах черепных нервов. Периферический отросток такого нейрона, имеющий чувствительное окончание — рецептор, берёт начало в органах и тканях. Центральный отросток в составе задних корешков спинномозговых нервов или чувствительных корешков черепных нервов направляется к соответствующим ядрам в спинной или головной мозг.
Второе звено рефлекторной дуги является эфферентным, поскольку несёт импульсы из спинного или головного мозга к рабочему органу. Этот эфферентный путь вегетативной рефлекторной дуги представлен двумя нейронами. Первый из этих нейронов, второй по счёту в простой вегетативной рефлекторной дуге, располагается в вегетативных ядрах ЦНС. Его можно называть вставочным, так как он находится между чувствительным (афферентным) звеном рефлекторной дуги и вторым (эфферентным) нейроном эфферентного пути.
Эффекторный нейрон представляет собой третий нейрон вегетативной рефлекторной дуги. Тела эффекторных (третьих) нейронов лежат в периферических узлах вегетативной нервной системы (симпатический ствол, вегетативные узлы черепных нервов, узлы внеорганных и внутриорганных вегетативных сплетений). Отростки этих нейронов направляются к органам и тканям в составе органных вегетативных или смешанных нервов. Заканчиваются постганглионарные нервные волокна на гладких мышцах, железа́х и в других тканях соответствующими концевыми нервными аппаратами.
Физиология
Общее значение вегетативной регуляции
Вегетативная нервная система приспосабливает работу внутренних органов к изменениям окружающей среды. ВНС обеспечивает гомеостаз (постоянство внутренней среды организма). ВНС также участвует во многих поведенческих актах, осуществляемых под управлением головного мозга, влияя не только на физическую, но и на психическую деятельность человека.
Роль симпатического и парасимпатического отделов
Симпатическая нервная система активируется при стрессовых реакциях. Для неё характерно генерализованное влияние, при этом симпатические волокна иннервируют подавляющее большинство органов.
Известно, что парасимпатическая стимуляция одних органов оказывает тормозное действие, а других — возбуждающее действие. В большинстве случаев действие парасимпатической и симпатической систем противоположно.
2.3. Нейромедиаторы и клеточные рецепторы
Симпатический и парасимпатический отделы оказывают различное, в ряде случаев противонаправленное влияние на различные органы и ткани, а также перекрёстно влияют друг на друга. Различное воздействие этих отделов на одни и те же клетки связано со спецификой выделяемых ими нейромедиаторов и со спецификой рецепторов, имеющихся на пресинаптических и постсинаптических мембранах нейронов автономной системы и их клеток-мишеней (Рис.).

Преганглионарные нейроны обоих отделов автономной системы в качестве основного нейромедиатора выделяют ацетилхолин, который действует на никотиновые рецепторы ацетилхолина на постсинаптической мембране постганглионарных (эффекторных) нейронов. Постганглионарные нейроны симпатического отдела, как правило, выделяют в качестве медиатора норадреналин, который действует на адренорецепторы клеток-мишеней. На клетках-мишенях симпатических нейронов бета-1 и альфа-1 адренорецепторы в основном сосредоточены на постсинаптических мембранах (это означает, что in vivo на них действует в основном норадреналин), а аль-2 и бета-2 рецепторы — на внесинаптических участках мембраны (на них в основном действует адреналин крови). Лишь некоторые постганглионарные нейроны симпатического отдела (например, действующие на потовые железы) выделяют ацетилхолин.
Постганглионарные нейроны парасимпатического отдела выделяют ацетилхолин, который действует на мускариновые рецепторы клеток-мишеней.

На пресинаптической мембране постганглионарных нейронов симпатического отдела преобладают два типа адренорецепторов: альфа-2 и бета-2 адренорецепторы. Кроме того, на мебране этих нейронов расположены рецепторы к пуриновым и пиримидиновым нуклеотидоам (P2X-рецепторы АТФ и др.), никотиновые и мускариновые холинорецепторы, рецепторы нейропептидов и простагландинов, опиоидные рецепторы[6].
При действии на альфа-2 адренорецепторы норадреналина или адреналина крови падает внутриклеточная концентрация ионов Ca2+, и выделение норадреналина в синапсах блокируется. Возникает петля отрицательной обратной связи. Альфа-2 рецепторы более чувствительны к норадреналину, чем к адреналину.
При действии норадреналина и адреналина на бета-2 адренорецепторы выделение норадреналина обычно усиливается. Этот эффект наблюдается при обычном взаимодействии с Gs-белком, при котором растёт внутриклеточная концентрация цАМФ. Бета-два рецепторы более чувствительны к адреналину. Поскольку под действием норадреналина симпатических нервов из мозгового слоя надпочечников выделяется адреналин, возникает петля положительной обратной связи.
Однако в некоторых случаях активация бета-2 рецепторов может блокировать выделение норадреналина. Показано, что это может быть следствием взаимодействия бета-2 рецепторов с Gi/o белками и связывания (секвестирования) ими Gs-белков, которое, в свою очередь, предотвращает взаимодействие Gs-белков с другими рецепторами [1].
При действии ацетилхолина на мускариновые рецепторы симпатических нейронов выделение норадреналина в их синапсах блокируется, а при действии на никотиновые рецепторы — стимулируется. Поскольку на пресинаптических мембранах симпатических нейронов преобладают мускариновые рецепторы, обычно активация парасимпатических нервов снижает уровень выделения норадреналина из симпатических нервов.
На пресинаптических мембранах постганглионарных нейронов парасимпатического отдела преобладают альфа-2 адренорецепторы. При действии на них норадреналина выделение ацетилхолина блокируется. Таким образом, симпатические и парасимпатические нервы взаимно ингибируют друг друга.
2.4. Регуляция тонуса гладких мышц сосудов
Схема. Биохимические механизмы сокращения и расслабления гладкомышечного миоцита. | |
 | |
Обозначения: | |
AC, adenylyl cyclase = аденилциклаза; Ca++кальция; DAG, diacylglycerol = диацилглицерол; Gs и Gα = белки G-рецепторы; IP3, inositol triphosphate = инозитолтрифосфат; LC20, light chain 20 - лёгкие цепи миозина 20; PIP3, phosphatidylinositol 3, 4, 5-triphosphate = фосфатидилинозитол 3,4,5-трифосфат; PLC, phospholipase = фосфолипаза; R-PKA, inactive protein kinase = инактивированная протеинкиназа. | |
2.5. Ненорадренергические и нехолинергические (NANC) вещества в регуляции сосудов
Кроме классических нейротрансмиттеров вегетативной нервной системы ацетилхолина и норадреналина, в литературе описаны многочисленные биологически активные вещества различного строения, модулирующие процессы передачи сигнала. К ним следует отнести серотонин (5-НТ), АТФ, кальцитонин ген связанный пептид (CGRP), допамин, ГАМК, Гонадотропин релизинг гормон (GnRH), нейропептид Y (NPY), вещество Р, вазоактивный кишечный пептид (VIP). Основные сведения о них собраны в таблице
|
Медиатор |
Тип |
Местоположение |
Функция |
|
Ганглии вегетативной нервной системы |
|||
|
АТФ |
постганглионарные симпатические нейроны (например, в кровеносных сосудах и семявыносящих протоках) |
Быстрая деполяризация/сокращение (вазоконстрикция) |
|
|
CGRP |
расширение кровеносных сосудов увеличение проницаемости сосудов |
||
|
Некоторые симпатические нервы (например, почки) |
|||
|
Ганглии вегетативной нервной системы |
|||
|
GnRH |
медленная деполяризация ацетилхолиновый котрансмиттер |
||
|
постганглионарные симпатические нейроны (например, кровеносные сосуды) |
Усиливают вазоконстрикторное действие норадреналина, ингибитор высвобождения норадреналина |
||
|
ацетилхолиновый котрансмиттер |
|||
|
парасимпатические нервы слюнных желез NANC иннервация гладких мышц дыхательных путей парасимпатические ганглии в сфинктерах, желчном пузыре и тонкой кишке |
расширение кровеносных сосудов ацетилхолиновый котрансмиттер расслабление кишечной гладкой мышцы и сфинктеров |
3.1. Ренин, ангиотензин, альдостероновая система (РААС или РАС)
Компоненты системы
Ангиотензиноген
Ангиотензин I
Ангиотензин II
Проренин
Ренин
Ангиотензинпревращающий фермент
Альдостерон
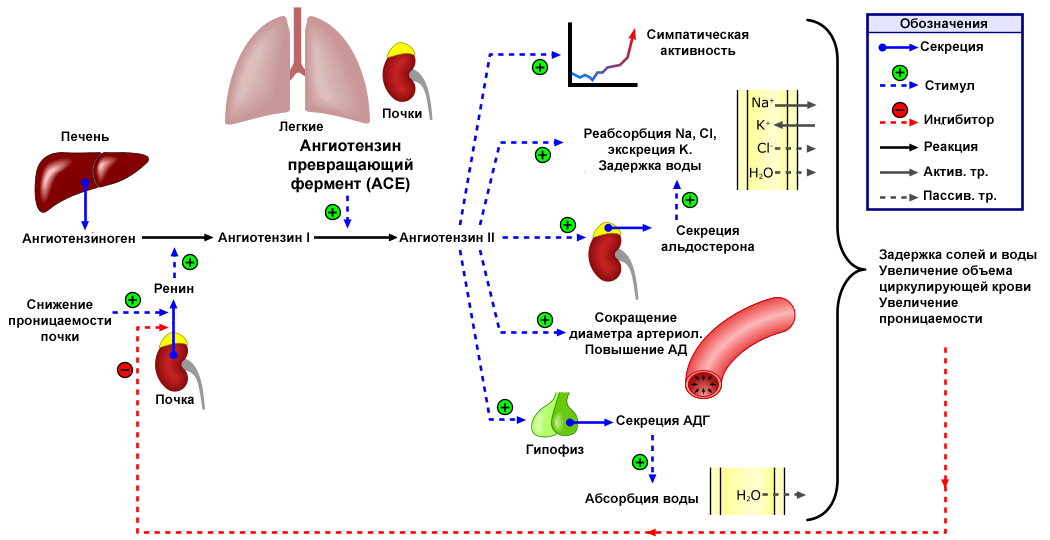
3.2. Продукция и секреция ренина
Ренин-ангиотензин альдестероновый каскад начинается с биосинтеза препрорениновой из рениновой мРНК в юкстагломерулярных клетках и превращается в проренин путём отщепления 23 аминокислот. В эндоплазматическом ретикулуме проренин подвергается гликозилированию и приобретает 3-D структуру, которая характерна для аспартатных протеаз. Готовая форма проренина состоит из последовательности включающей 43 остатка присоединённых к N-концу ренина, содержащего 339-341 остаток. Предполагается, что дополнительная последовательность проренина (prosegment) связана с ренином для предотвращения взаимодействия с ангиотензиногеном. Большая часть проренина свободно выбрасывается в системный кровоток путём экзоцитоза, но некоторая доля превращается в ренин путём действия эндопептидаз в секреторных гранулах юкстагломерулярных клеток. Ренин, образуемый в секреторных гранулах в дальнейшем выделяется в кровоток, но этот процесс жёстко контролируется давлением, ангиотензином 2, NaCl, через внутриклеточные концентрации ионов кальция. Поэтому у здоровых людей объём циркулирующего проренина в десять раз выше концентрации активного ренина в плазме . Однако, все же остаётся не понятным, почему концентрация неактивного предшественника настолько высока.
Контроль секреции ренина
Активная секреция ренина регулируется четырьмя независимыми факторами:
- Почечным барорецепторным механизмом в афферентной артериоле, который улавливает изменение почечного перфузионного давления.
- Изменениями уровня NaCl в дистальном отделе нефрона. Этот поток измеряется как изменение концентрации Cl- клетками плотного пятна дистального извитого канальца нефрона в области, прилегающей к почечному тельцу.
- Стимуляцией симпатическими нервами через бета-1 адренергические рецепторы.
- Механизмом отрицательной обратной связи, реализованным через прямое действие ангиотензина 2 на юкстагломерулярные клетки.
Секрецию ренина активирует снижение перфузионного давления или уровня NaCl и повышение симпатической активности. Ренин также синтезируется и в других тканях, включая мозг, надпочечник, яичники, жировая ткань, сердце и сосудах.
Контроль секреции ренина — определяющий фактор активности РААС.
3.3. Ангиотензиноген, ангиотензины 1,2,3,4 и т.д.
Ренин регулирует начальный, ограничивающий скорость, этап РААС путём отщепления N-концевого сегмента ангиотензиногена для формирования биологически инертного декапептида ангиотензина 1 или Ang-(1-10). Первичный источник ангиотензиногена — печень. Долговременный подъём уровня ангиотензиногена в крови, который происходит во время беременности, при синдроме Иценко-Кушинга или при лечении глюкокортикоидами, может вызвать гипертензию, хотя и существуют данные о том, что хроническое повышение концентрации ангиотензина в плазме частично компенсируется снижением секреции ренина. Неактивный декапептид Ang 1 гидролизуется ангиотензинпревращающим ферментом (АПФ), который отщепляет С-концевой дипептид и, таким образом, формируется октапептид Ang 2 [Ang-(1-8)], биологически активный, мощный вазоконстриктор. АПФ представляет собой экзопептидазу и секретируется главным образом лёгочным и почечным эндотелием, нейроэпителиальными клетками. Ферментативная активность АПФ заключается в повышении вазоконстрикции и снижении вазодилятации.
Хотя Ang2 наиболее биологически активный продукт РААС, существуют данные, что другие метаболиты агиотензинов 1 и 2 могу также могут иметь значительную активность. Ангиотензин 3 и 4 (Ang 3 & Ang 4) формируются путём отщепления аминокислот с N-конца от Ангиотензина 2 вследствие действия аминопептидаз А и N. Ang 3 и 4 наиболее часто вырабатываются в тканях с высоким содержанием этих ферментов, например, в мозге и почках. Ang 3 [Ang-(2-8)], гептапептид образующий в результате отщепления аминокислоты с N-конца, наиболее часто он встречается в центральной нервной системе, где Ang III играет важную роль в поддержании кров давления. Ang IV [Ang-(3-8)] гексапептид является результатом дальнейшего ферментативного расщепления AngIII. Предполагается, что Ang 2 и 4 работают кооперативно. В качестве примера можно привести повышение кровяного давления в мозгу, вызываемое действием этих ангиотензинов на AT1-рецептор. Причём этот гемодинамический эффект Ang 4 требует наличия как Ang2 так и самого AT1- рецептора. Пептиды, получаемые отщеплением аминокислот с С-конца, могут также иметь биологическую активность. Например, Ang-(1-7), гептапептидный фрагмент ангиотензина 2, может образовываться как из Ang2 так и из Ang1 действием ряда эндопептидаз или действием карбоксипептидаз (например, гомологом АПФ, названным АПФ2) конкретно на Ang2. В отличие от АПФ, АПФ2 не может участвовать в реакции превращения Ang1 в Ang2 и его активность не подавляется ингибиторами ACE (ACEIs). Ang-(1-7) реализующий свои функции через определённые рецепторы, впервые был описан как вазодилататор и как натуральный ингибитор ACEI. Ему также приписываются и кардиопротекторные свойства. АПФ2 может также отщеплять одну аминокислоту с С-конца, результатом такого действия является Ang-(1-9), пептид с неизвестными функциями.
3.4. Реакции, катализируемые АПФ
Превращение неактивного ангиотензина I (представляет собой декапептид – последовательность из 10 аминокислот Asp-Arg-Val-Tyr-Ile-His-Pro-Phe-His-Leu) в активный октапептид ангиотензин II (путем удаления 2 аминокислот-His-Leu) контролирует ангиотензинпревращающий фермент (АПФ) – циркулирующий во внеклеточном пространстве белок (карбоксипептидаза). Вторая важная функция АПФ – деактивация брадикинина.
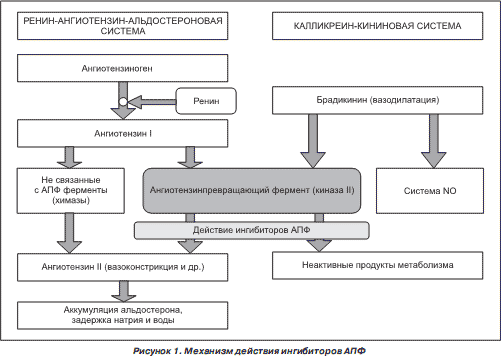
Ген ACE порождает два изофермента АПФ — соматический, обнаруживаемый во многих органах и тканях, и герминальный, присутствующий только в сперме.
АПФ типичный секреторный фермент. Кроме регуляции кровяного давления, АПФ участвует в различных процессах, происходящих в организме. Его синтезируют клетки многих тканей, таких как васкулярные эндотелиальные клетки, почечные эпителиальные клетки, тестикулярные клетки Лейдига и т. д.
3.5. Уровень АПФ в крови и особенности генетики человека
В норме у разных людей уровень АПФ в плазме крови может различаться до 5 раз. У конкретного же человека уровень АПФ достаточно стабилен. Такие колебания уровня АПФ между людьми вызваны полиморфизмом гена АСЕ. В 16-м интроне гена выявлен инсерционно-делеционный (I/D) полиморфизм, заключающийся во вставке (инсерции, I) или потере (делеции, D) Alu-повтора, размером в 289 пар нуклеотидов. Делеция Alu-повтора приводит к повышению экспрессии гена ACE и увеличению концентрации АПФ в крови, лимфе и тканях, что является фактором, повышающим риск развития сердечно-сосудистых заболеваний (инфаркта миокарда, гипертрофии левого желудочка, ишемической болезни сердца), болезни почек, атеросклероза, болезни Альцгеймера.
У лиц, гомозиготных по аллелю D (генотип D/D), уровень АПФ повышен в 2 раза по сравнению с I/I генотипом. Ассоциация D/D-генотипа с гипертрофией левого желудочка сильнее у мужчин, чем у женщин, таким образом, инсерционно-делеционный полиморфизм может выступать как маркер риска развития гипертрофии левого желудочка у мужчин среднего возраста.
Выявлена ассоциация полиморфизма с возрастной макулярной дегенерацией (ВМД) – основной причиной потери зрения в пожилом возрасте. Генотип I/I встречается в 4,5 раза чаще у здоровых людей, чем у пациентов, страдающих ВМД, и, по-видимому, защищает от ВМД.
Ассоциации полиморфизма АСЕ с уровнем систолического и диастолического кровяного давления не выявлено, кроме случаев злокачественной формы гипертонической болезни (быстро прогрессирующая и тяжело протекающая, со значительным повреждением сосудов почек и сетчатки), при которой генотип D/D встречается чаще.
Также D-аллель связан с риском развития нефропатии у больных сахарным диабетом.
Аллель I ассоциирован с повышенной устойчивостью организма к физическим нагрузкам. Низкая минеральная плотность костной ткани и мышечная слабость являются основными факторами риска переломов костей у женщин при остеопорозе в постменопаузе. Выявлено, что наличие в генотипе женщины аллеля I положительно влияет на эффективность тренировки мышц (у пациенток с генотипом I/I она оказалась в 2 раза выше, чем у женщин с генотипом D/D) в ответ на физические нагрузки, на фоне гормональной терапии.
Исследования также показывают, что у высококлассных спортсменов-стайеров увеличена частота аллеля I, ассоциированная с выносливостью.
3.6. АПФ как мишень фармакологического воздействия
В 1991 году японские ученые создали первый ингибитор АПФ на молочной основе, в виде ферментированного молочного напитка, с использованием конкретных культур, чтобы освободить трипептид изолейцин-пролин-пролина (IPP) из молочного белка. Валин-пролин-пролин (VPP) также выделяется в этом процессе-другое молоко трипептиде с очень схожей химической структурой в IPP. Вместе эти пептиды теперь часто называют lactotripeptides. В 1996 году первое исследование человеческого подтвердили артериальное давление эффект снижения IPP в ферментированного молока. [57] Хотя в два раза количество VPP необходимо, чтобы достигнуть того же самого АСЕ-ингибирующей активности, как первоначально обнаружена IPP, VPP также предполагается, чтобы добавить к общему эффекту снижения кровяного давления. [58] С момента открытия первых lactotripeptides, более 20 клинических испытаний человека были проведены в разных странах
3.7. Классификация ингибиторов АПФ
Классификация ингибиторов АПФ
Препараты, содержащие сульфгидрильные группы: каптоприл, зофеноприл.
Дикарбоксилат-содержащие препараты: эналаприл, рамиприл, хинаприл, периндоприл, лизиноприл, беназеприл.
Фосфонат-содержащие препараты: фозиноприл.
Природные ингибиторы АПФ.
3.8. Функциональная активность ангиотензина II
Сердечно-сосудистая система
Ангиотензин II — сильный вазоконстриктор прямого действия. Он сужает артерии и вены, что приводит к повышению давления. Сосудосуживающая активность ангиотензина II определяется его взаимодействием с AT1 рецептором. Лиганд-рецепторный комплекс активирует НАДФH-оксидазу, образующую супероксид, который в свою очередь взаимодействует с вазорелаксирующим фактором оксидом азота NO и инактивирует его. Кроме этого, он обладает протромботическим эффектом, регулируя адгезию и агрегацию тромбоцитов и синтез ингибиторов PAI-1 и PAI-2. К отсроченным эффектам АТII гипертрофию мышечного слоя сосудов и гипертрофию левого желудочка миокарда.
Нервная система
Ангиотензин вызывает чувство жажды. Он повышает секрецию антидиуретического гормона в нейросекреторных клетках гипоталамуса и секрецию АКТГ в передней доле гипофиза, а также потенциирует высвобождение норадреналина за счёт прямого действия на постганглионарные симпатические нервные волокна.
Надпочечники
Под действием ангиотензина кора надпочечников выделяет гормон альдостерон, вызывающий задержку натрия и потерю калия.
Почки
Ангиотензин обладает прямым эффектом на проксимальные канальцы, что увеличивает задержку натрия. В целом ангиотензин увеличивает скорость гломерулярной фильтрации за счёт сужения эфферентных почечных артериол и повышения давления в почках.
3.9. Ангиотензиновые рецепторы
Ангиотензиновые рецепторы - G-белоксопряжённые рецепторы, лигандами которых является ангиотензин, гормон ренин-ангиотензиновой системы, и его производные. У человека описано четыре AT-рецептора. Антагонисты рецепторов ангиотензина II применяются в качестве средств, снижающих давление.
| Подтип рецептора | Внутриклеточный механизм |
|---|---|
| AT1 | |
| AT2 | |
| AT3 | |
| AT4 |
Рецептор АТ1 является наиболее изученным.
Локализация. АТ1 подтип находится в сердце, кровеносные сосуды, почки, надпочечники коры головного мозга, легкое и головной мозг и опосредует сосудосуживающие эффекты. Молекулярный механизм. Активированный рецептор освобождает Gq/11 и Gi/о и тем самым активирует фосфолипазу С и повышает концентрации цитозольного Ca2+, который, в свою очередь, запускает клеточные ответы, такие как стимуляция протеинкиназы С. Активированный рецептор также ингибирует аденилатциклазу и активирует различные тирозинкиназы.
Эффекты, опосредованные АТ1 рецептора включают вазоконстрикцию, повышение секреции и синтеза альдостерона, повышение секреции вазопрессина, гипертрофию сердца, увеличение периферической норадренергической активности, сосудистой пролиферации клеток гладких мышц, снижение почечного кровотока, ингибирование высвобождения ренина, обратного захвата натрия в почечных канальцах, модуляция центральной активности симпатической нервной системы, сердечной сократимости, центрального контроля за осмотическим давлением и формирования внеклеточного матрикса.
Основные эффекты стимуляции различных типов рецепторов ангиотензина II
| Рецепторы 1-го типа | Рецепторы 2-го типа |
Вазоконстрикция | Вазодилатация |
АТ2-рецепторы более многочисленны у плода и новорожденного. Знания о рецепторе АТ2 остаются загадочными и противоречивыми - он, вероятно, участвуют в сосудистом росте. Эффекты, опосредованные АТ2-рецептора предлагается, чтобы включать в себя ингибирование роста клеток, развитие ткани плода, модуляцию внеклеточного матрикса, регенерацию нейронов, апоптоза, клеточной дифференцировки, и, возможно, вазодилатацию и гипертрофию левого желудочка.
АТ3 и АТ4 плохо охарактеризованые подтипы. Рецепторы АТ4 активируются метаболитом ангиотензина II ангиотензином IV, и могут играть роль в регуляции внеклеточного матрикса ЦНС, а также модуляции высвобождения окситоцина
3.10. Ангиотензиновые рецепторы как мишень фармакологического воздействия.
Антагонисты рецепторов ангиотензина (АРА) — относительно новый класс среди гипотензивных препаратов первого ряда, рекомендованных для лечения артериальной гипертензии ВОЗ/МОГ в 1999 году. Первый препарат АРА для перорального применения — лосартан — был синтезирован группой исследователей во главе с P.Timmermans в 1988 году и применяется в клинике с 1994 года. В целом АРА оказывают антигипертензивный эффект, аналогичный другим препаратам первого ряда, но отличаются низкой частотой побочных эффектов и очень хорошей переносимостью.
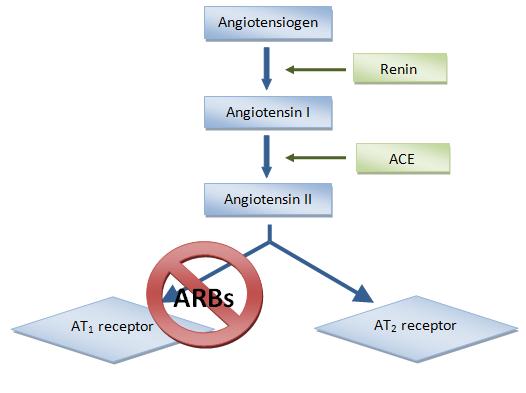
Препараты этой группы не влияют на образование ангиотензина II. Точка приложения их действия —рецепторы ангиотензина II 1-го типа — находится на периферии биохимического каскада ренин-ангиотензиновой системы (см. рис. 9.1). Избирательное действие АРА именно на рецепторы 1 -го типа ангиотензина II позволяет устранить негативные влияния РАС на сердечно-сосудистую систему, вместе с нежелательными эк- стракардиальными эффектами. В то же время сохраняются потенциально положительные эффекты стимуляции рецепторов 2-го типа (антипролиферативное действие, вазодилатация).

В 1970-х годах было показано, что повышение уровня Ang II может нанести вред сердцу и почкам, а люди с высоким уровнем активности ренина в плазме подвергались повышенному риску инфаркта миокарда и инсульта [4]. С введением ингибиторов ангиотензинпревращающего фермента (АПФ) в конце 1970-х годов было подтверждено, что Ang II играет важную роль в регулировании артериального давления, электролитов и баланса жидкости [5].
Ang II связывается с АТ1-рецепторами через различные сайты связывания [1]. Первичный сайт связывания находится во внеклеточной области АТ1-рецептора, где Ang II взаимодействует с остатками в N-конце рецептора AT1 и его первой и третьей внеклеточных петель. Трансмембранные спирали также способствуют связыванию через С-концевую карбоксильную группу, которая взаимодействует с Lys199 в верхней части спирали 5 рецептора, подробности см. На рисунке 1. [8]

Ионный мост, образованный между Lys199 и карбоксильной концевой группой остатка Phe8 Ang II, скорее всего, стабилизирован остатком Trp253. Кроме того, Phe259 и Asp263 в трансмембранной спирали 6 и Lys102 и Ser105 во внешней области трансмембранной спирали 3 также были вовлечены в связывание Ang II. Эта область может, возможно, участвовать в стабилизации ратификации рецептора и в образовании внутримембранного связующего кармана.

Большинство АРБ имеют один и тот же фармакофор, поэтому разница в их биохимическом и физиологическом эффектах в основном обусловлена различными заместителями. Активность лекарственного средства зависит от его сродства к сайту субстрата и продолжительности его связывания с сайтом. Липофильные заместители, такие как линейная алкильная группа в 2-положении на имидазольном кольце вместе с бифенилметильной группой, связываются с гидрофобными карманами рецептора. Кислотная группа, такая как тетразол, CO2H или NHSO2CF3 в 1-положении бифенилметильной группы, будет связываться с основным положением в рецепторе и необходима для сильной антагонистической активности [18].
В валсартане имидазольное кольцо лозартана было заменено ацилированной аминокислотой [4].
Несколько заместителей были испытаны в 4- и 5- положениях на имидазольном кольце. Хлор и гидроксиметильные группы, связанные с этими положениями в лозартане, вероятно, не имеют большого значения при связывании рецепторов, поскольку другие АРБ не обладают этими функциональными группами и имеют сравнимые или лучшие связывающие аффинности, чем лозартан. Ирбезартан имеет карбонильную группу в 5-положении, функционирующую в качестве акцептора водородной связи вместо гидроксиметильной группы лозартана, что приводит к более длительному связыванию с рецептором [1] [4] [18] Структура eprosartan отличается тем, что отличается от других ARB, обычная бифенилметильная группа была заменена карбоксибензильной группой, которая более точно имитирует фенольную группу Tyr4 группы Ang II. Это изменение приводит к более сильному связыванию с рецептором, но биохимические и физиологические эффекты существенно не улучшаются [1].
Telmisartan имеет карбоновую кислоту в 2-положении бифенилметильной группы и является более сильным, чем аналог тетразола [1]. Сообщалось, что имидазолы, которые имеют гидроксиметил- и карбоксильные группы в положении 4 и 5, обладают сильной антагонистической активностью, вызванной водородной связью и гидрофильностью гидроксиметильной группы [18]. Сообщалось также, что гидроксильная группа в 4-положении на имидазольном кольце играет важную роль в аффинности связывания и компенсирует недостаток липофильности объемной алкильной группы. [18] Эти результаты показывают, что гидроксиалкильная группа среднего размера, такая как CHMeOH и CMe2OH, благоприятна для заместителя 4-позиции на имидазольном кольце. Кроме того, ионизируемая группа благоприятна для аффинности связывания [18].
Большинство АРБ имеют один и тот же фармакофор, поэтому разница в их биохимическом и физиологическом эффектах в основном обусловлена различными заместителями. Активность лекарственного средства зависит от его сродства к сайту субстрата и продолжительности его связывания с сайтом. Липофильные заместители, такие как линейная алкильная группа в 2-положении на имидазольном кольце вместе с бифенилметильной группой, связываются с гидрофобными карманами рецептора. Кислотная группа, такая как тетразол, CO2H или NHSO2CF3 в 1-положении бифенилметильной группы, будет связываться с основным положением в рецепторе и необходима для сильной антагонистической активности [18].
В валсартане имидазольное кольцо лозартана было заменено ацилированной аминокислотой [4].
Несколько заместителей были испытаны в 4- и 5- положениях на имидазольном кольце. Хлор и гидроксиметильные группы, связанные с этими положениями в лозартане, вероятно, не имеют большого значения при связывании рецепторов, поскольку другие АРБ не обладают этими функциональными группами и имеют сравнимые или лучшие связывающие аффинности, чем лозартан. Ирбезартан имеет карбонильную группу в 5-положении, функционирующую в качестве акцептора водородной связи вместо гидроксиметильной группы лозартана, что приводит к более длительному связыванию с рецептором [1] [4] [18] Структура eprosartan отличается тем, что отличается от других ARB, обычная бифенилметильная группа была заменена карбоксибензильной группой, которая более точно имитирует фенольную группу Tyr4 группы Ang II. Это изменение приводит к более сильному связыванию с рецептором, но биохимические и физиологические эффекты существенно не улучшаются [1].
Telmisartan имеет карбоновую кислоту в 2-положении бифенилметильной группы и является более сильным, чем аналог тетразола [1]. Сообщалось, что имидазолы, которые имеют гидроксиметил- и карбоксильные группы в положении 4 и 5, обладают сильной антагонистической активностью, вызванной водородной связью и гидрофильностью гидроксиметильной группы [18]. Сообщалось также, что гидроксильная группа в 4-положении на имидазольном кольце играет важную роль в аффинности связывания и компенсирует недостаток липофильности объемной алкильной группы. [18] Эти результаты показывают, что гидроксиалкильная группа среднего размера, такая как CHMeOH и CMe2OH, благоприятна для заместителя 4-позиции на имидазольном кольце. Кроме того, ионизируемая группа благоприятна для аффинности связывания [18].
Candesartan и olmesartan имеют самое высокое сродство к рецепторам AT1, за которыми следуют ирбесартан и эпросартан. Валсартан, телмисартан и EXP 3174 имеют сходные сродства, которые примерно в десять раз меньше, чем у кандесартана. Лозартан имеет наименьшее сходство. [6] Сродство ARB к рецептору AT2 обычно значительно ниже (или примерно в 10 000 раз меньше), чем для подтипа AT1. Поэтому они допускают беспрепятственную стимуляцию АТ2-рецептора [19].
Candesartan и olmesartan имеют самое высокое сродство к рецепторам AT1, за которыми следуют ирбесартан и эпросартан. Валсартан, телмисартан и EXP 3174 имеют сходные сродства, которые примерно в десять раз меньше, чем у кандесартана. Лозартан имеет наименьшее сходство. [6] Сродство ARB к рецептору AT2 обычно значительно ниже (или примерно в 10 000 раз меньше), чем для подтипа AT1. Поэтому они допускают беспрепятственную стимуляцию АТ2-рецептора [19].
3.11. Неприлизин
К вазопептидазам кроме известного АПФ относятся еще 2 других цинк-металлопротеиназы – неприлизин (нейтральная эндопептидаза, НЭП) и эндотелин-превращающий фермент, которые также могут быть мишенями для фармакологического воздействия.
Неприлизин, также известный как мембранная металлоэндопептидаза (MME), нейтральная эндопептидаза (NEP), кластер дифференцировки 10 (CD10) и общий острый лимфобластный лейкозный антиген (CALLA) - это фермент, который у человека кодируется геном MME. Неприлизин представляет собой цинк-зависимую металлопротеазу, которая расщепляет пептиды на аминогруппе гидрофобных остатков и инактивирует несколько пептидных гормонов, включая глюкагон, энкефалины, вещество Р, нейротензин, окситоцин и брадикинин. Он также деградирует бета-пептид амилоида, аномальное расстройство и агрегация в нейронной ткани которого являются причиной болезни Альцгеймера. Синтезированный в виде белка, связанного с мембраной, эктодомен неприлизина высвобождается во внеклеточный домен после того, как он был перенесен с аппарата Гольджи на поверхность клетки.

Система натрийуретического пептида представлена тремя разными изоформами: предсердным натрийуретическим пептидом (А-тип), мозговым натрийуретическим пептидом (В-тип), которые синтезируются в предсердии и миокарде, и эндотелиальным С-пептидом, которые по своим биологическим функциям являются эндогенными ингибиторами РААС и эндотелина-1 (табл. 1) [3, 4]. Кардиоваскулярные и ренальные эффекты натрийуретического пептида заключаются в снижении АД через влияние на сосудистый тонус и водноэлектролитный баланс, а также в антипролиферативном и антифибротическом действии на органы-мишени. По самым последним данным, система натрийуретического пептида участвует в метаболической регуляции: окислении липидов, образовании и дифференцировке адипоцитов, активации адипонектина, секреции инсулина и толерантности к углеводам, что может обеспечивать защиту от развития метаболического синдрома.
К настоящему времени стало известно, что развитие сердечно-сосудистых заболеваний ассоциируется с дизрегуляцией системы натрийуретического пептида. Так, при АГ наблюдается дефицит натрийуретического пептида, приводящий к солечувствительности и нарушению натрийуреза; при хронической сердечной недостаточности (ХСН) на фоне дефицита наблюдается аномалия функционирования гормонов системы натрийуретического пептида.
Поэтому для потенцирования системы натрийуретического пептида с целью достижения дополнительного гипотензивного и протективных кардиоренальных эффектов возможно применение ингибиторов НЭП. Ингибирование неприлизина приводит к потенцированию натрийуретического, диуретического и вазодилатирующего эффектов эндогенного натрийуретического пептида и в результате – к снижению АД. Однако НЭП участвует в деградации и других вазоактивных пептидов, в частности АТI, АТII и эндотелина-1. Поэтому баланс эффектов воздействия на сосудистый тонус ингибиторов НЭП вариабельный и зависит от преобладания констрикторных и дилатирующих влияний. При длительном применении антигипертензивное действие ингибиторов неприлизина выражено слабо вследствие компенсаторной активации образования АТII и эндотелина-1.
В этой связи сочетание эффектов ингибиторов АПФ и ингибиторов НЭП может существенно потенцировать гемодинамические и антипролиферативные эффекты в результате комплементарного механизма действия, что привело к созданию препаратов с двойным механизмом действия, объединенных названием – ингибиторы вазопептидаз
3. Corti R., Burnett J.C., Rouleau J.L. et al. Vasopeptidase inhibitors: a new therapeutic concept in cardiovascular disease? // Circulation. 2001. Vol. 104. P. 1856–1862.
4. Mangiafico S., Costello-Boerrigter L.C., Andersen I.A. et al. Neutral endopeptidase inhibition and the natriuretic peptide system: an evolving strategy in cardiovascular therapeutics // Eur. Heart J. 2012, doi:10.1093/eurheartj/ehs262.
4. Эндотелий-зависимые факторы
Эндотелий выполняет ключевую функцию в регуляции тонуса и роста сосудов. Эндотелий сосудов обладает способностью синтезировать и выделять факторы, вызывающие расслабление или сокращение гладких мышц сосудов в ответ на разного рода стимулы (табл). Общая масса эндотелиоцитов, монослойно выстилающих кровеносные сосуды изнутри (интима), у человека приближается к 500 г. Общая масса, высокая секреторная способность эндотелиальных клеток позволяют рассматривать эту «ткань» как своеобразный эндокринный орган (железу). Распределенный по сосудистой системе эндотелий, очевидно, предназначен для паракринной регуляции гладкомышечных образований сосудов. Период полужизни выделяемого эндотелиоцитами инкрета очень мал. Для NO он составляет — 6—25 с (вследствие быстрого перехода его в нитраты и нитриты), но он способен сокращать и расслаблять гладкие мышцы сосудов, не оказывая влияния на эффекторные образования других органов (кишечник, бронхи, матка).
Таблица Некоторые биологические регуляторы, секретируемые эндотелием
|
Вазоконстрикторы |
Вазодилятаторы |
|
Эндотелин 1 |
NO |
|
Тромбоксан А2 |
Простациклин |
|
20-HETE (20-гидроксиэйкозотетраеновая кислота)о) |
Натрийуретический пептид С |
|
|
Кинины |
4.1. Эндотелин
Эндотелин — сосудосуживающий пептид из 21 аминокислоты, играющий ключевую роль в гомеостазе кровеносных сосудов. Эндотелин является самым мощным из известных сосудосуживающих агентов. Он в 10 раз более активен, чем ангиотензин II.
Эндотелий секретирует большой эндотелин (проэндотелин) (38 аминокислотных остатков). Под влиянием эндотелинпревращаюшего фермента, находящегося внутри и на поверхности эндотелия, из большого эндотелина образуются три изомера эндотелинов.
Эндотелины — бициклические полипептиды, состоящие из 21 аминокислотного остатка с двумя бисульфидными связями. Имеется большое сходство между структурой эндотелинов и некоторыми нейротоксическими пептидами.
Основной механизм действия эндотелинов заключается в высвобождении кальция, что вызывает:
- стимуляцию всех фаз гемостаза, начиная с агрегации тромбоцитов и заканчивая образованием красного тромба;
- сокращение и рост гладких мышц сосудов, приводящие к утолщению стенки сосудов и уменьшению их диаметра — вазоконстрикции.
Синтез эндотелинов усиливают тромбин (активизирующий эндотелинпревращающий фермент) и тромбоциты. Эндотелины, в свою очередь, вызывают адгезию и агрегацию тромбоцитов.
Существует 3 изоформы пептида, которые отличаются по спектру экспрессирующих их тканей, и 2 рецептора пептида ETA и ETB.
ETA рецептор находится в гладкомышечной ткани и при связывании с ним эндотелина вызывает сужение сосудов и задержку натрия, что повышает кровяное давление.
ETB рецептор находится на клетках эндотелия. Когда эндотелин связывается с ETB, это увеличивает вывод натрия и диурез (мочевыведение), а также приводит к повышению синтеза NO (или англ. endothelium-derived relaxing factor), обладающего сосудорасширяющими свойствами. Всё это снижает кровяное давление.
Эффекты эндотелинов неоднозначны и определяются рядом причин. Наиболее активен изомер — эндотелин-1. Он образуется не только в эндотелии, но и в гладких мышцах сосудов, нейронах, глие, мезенгиальных клетках почек, печени и других органах. Полупериод жизни — 10-20 мин., в плазме крови — 4-7 мин. Легкие удаляют до 90 % эндотелинов. Эндотелин-1 причастен к ряду патологических процессов (инфаркту миокарда, нарушению ритма сердца, легочной и системной гипертонии, атеросклерозу и др.).
Конечный эффект эндотелина зависит от его концентрации. При низкой концентрации больше проявляется его сосудорасширяющий эффект благодаря связыванию с ETB рецепторами на эндотелии. Однако, при более высокой концентрации он начинает связываться с ETA рецепторами на гладкомышечных клетках и проявляет свою основную активность, вызывая значительное сужение сосудов.
В норме существует баланс между сосудосуживающими (эндотелин, кальцитонин и др.) и сосудорасширяющими (NO, простациклин и др.) агентами. Такой баланс поддерживает и регулирует кровяное давление.
4.2. Старые лекарства для лечения гипертонии весьма эффективны, в то время как новые не являются достаточно инновационными
Вот краткое и простое руководство по улучшению понимания пяти наиболее распространенных препаратов, используемых большинством пациентов.
- Наиболее часто используемым препаратом является гидрохлоротиазид или HCTZ (который используется 22 процентами пациентов с гипертензией, согласно QuintilesIMS). Это мочегонное средство, которое заставляет почки увеличивать мочеиспускание. Это приводит к уменьшению объема жидкости в кровотоке, что снижает кровяное давление. Это или его кузина наркотиков, хлорталидон, являются предпочтительными первых препаратов, потому что они хорошо работают для большинства пациентов.
- Равно распространенным является лизиноприл, препарат в составе ингибитора ангиотензинпревращающего фермента (ACEI) (большинство из них заканчивается на «-приле»). Это мешает химическому сигналу в организме, который в противном случае повышал бы кровяное давление.
- Амлодипин, который используется 17 процентами пациентов с гипертонией, является блокатором кальциевых каналов. Он снижает кровяное давление, частично предотвращая попадание кальция в клетки, что препятствует сужению сосудов и повышению артериального давления.
- Класс лекарственного средства, тесно связанный с ACEI, представляет собой блокаторы рецепторов ангиотензина или ARB, которые препятствуют той же цепи реакций, что и ACEI. Наиболее часто используемый АРБ, лозартан, принимается 10% пациентов. Обычно ACEI назначаются первыми из-за их хорошо установленной записи, но заменяются ARB, если возникают побочные эффекты.
- В отличие от других классов, бета-блокаторы не являются отличным лекарством от артериального давления для всех. Они лучше всего подходят для людей с сердечными заболеваниями, потому что они замедляют сердечный ритм и снижают кровяное давление. Метопролол, используемый 8 процентами пациентов с гипертонией, является ведущим бета-блокатором (большинство заканчивается в «-olol»).
HCTZ, лизиноприл и амлодипин являются эффективными лекарственными средствами, которые часто используются вместе. Большинство людей требуют двух или трех разных лекарств, чтобы полностью контролировать кровяное давление (точно так же, как Маргарет). Иногда пациенты требуют еще больше лекарств.
Три лучших лекарств являются дешевыми лекарствами пожилого возраста. HCTZ, впервые доступный в 1959 году, стоит аптеках всего лишь в 4,29 доллара США за годовую поставку одного пациента. Lisinopril (введен в 1987 году и стоит аптек $ 6.33 в год на одного пациента) и амлодипин (введенный в 1992 году и стоимость аптек $ 5,66 в год на одного пациента) также недороги.
Есть новые наркотики, но они не прорывы. Многие напоминают существующие лекарства или представляют собой просто комбинации уже имеющихся лекарств. Для большинства пациентов они не стоят значительных дополнительных затрат. Например, azilsartan, новый блокатор рецепторов ангиотензина, введенный в 2011 году, стоит 2 000 долларов США в год на одного пациента.
Это отсутствие инноваций со стороны фармацевтических компаний связано с учетом огромного бремени болезней, вызванного неадекватным лечением высокого кровяного давления. На данный момент более старые препараты являются лучшими препаратами, но очень часто их нужно сочетать
Randall Stafford, MD, PhD, a professor of medicine and director of the Program on Prevention Outcomes and Practices, practices primary care internal medicine at Stanford.
http://scopeblog.stanford.edu/2017/10/12/too-high-older-drugs-work-well-for-hypertension-new-medications-show-little-innovation/