Введение в нейробиологию
4. Липиды мозга
Нейробиолог Филипп Хайтович: «Человеком нас сделал жир в мозге»
Свойства липидов мозга
- Липиды имеют множество функций в мозге.
- Мембранные липиды являются амфифильными молекулами.
- Гидрофобные компоненты многих липидов состоят либо из изопреноидов, либо из жирных кислот и их производных.
- Изопреноиды представляют собой структуры с пятью углеродными атомами.
- Жирные кислоты представляют собой карбоновые кислоты с длинной цепью, которые могут содержать одну или несколько двойных связей
Липиды являются не только структурными компонентами ЦНС, но и важнейшими участниками функциональной активности. Головной мозг характеризуется высоким содержанием липидов (более 50% сухой массы).
Мозг содержит уникальные мембранные структуры — миелиновые оболочки, которые имеют самое высокое содержание липидов (до 80%) по сравнению с другими тканями или субклеточными структурами, за исключением адипозной ткани. Для ЦНС характерно и наибольшее структурное разнообразие липвдов по сравнению с мембранами других органов. Липидный состав нервной ткани практически постоянен и остается неизменным даже под влиянием внешних факторов (диета, гормоны, фармакологические вещества, стрессы), которые меняют липидный состав висцеральных органов и плазмы. Это — следствие защищенности ЦНС от различных внешних воздействий. Изменение липидного состава нервной ткани рассматривается обычно как патология, хотя при этом следует помнить, что существенные изменения в липидном составе нервной системы происходят в период развития.
Вся сложнейшая деятельность нервной ткани опосредуется через мембраны, в формировании и функционировании которых липиды принимают непосредственное участие.
В
клетках нервной системы представлено несколько типов высокоспециализированных
мембран: соматические мембраны мульти- и униполярных нейронов, мембраны
дендритов, миелинизированных и немиелинизированных аксонов, аксонного холмика,
где генерируется потенциал действия, мембраны рыхлого и компактного миелина,
мембраны синаптических пузырьков, пре- и постсинаптические мембраны, мембраны
макро- и микроглии. Возбудимость этих мембран колеблется в широких пределах от
высоковозбудимых (синаптические, аксонного холмика) до относительно устойчивых
мультимембранных структур миелина. В составе, строении и функционировании
мембран нервной ткани еще очень много неясного. Для того чтобы раскрыть
надмолекулярную организацию этих мембран, надо иметь достаточно полное
представление об их липидном и белковом составе. Однако исследователи пока не
владеют этими сведениями в полной мере , хотя ряд важных закономерностей уже
намечен.
Липидный состав серого и белого вещества мозга человека представлен в табл.
4.1, а различных клеток мозга — в табл. 4.2. Видно, что липидный состав белого
вещества ближе к миелину, а серое вещество содержит меньше типичных миелиновых
липидов (цереброзидов, сульфатидов, фосфатидилэтаноламина), но относительно
больше ганглиозидов.
Сравнивая молярное содержание основных классов липидов в специализированных клетках мозга, можно видеть, что олигодендроглия и миелин наиболее обогащены цереброзидами, а нейроны и астроглия имеют более высокое содержание фосфолипидов. Это лишний раз подтверждает, что плазматические мембраны совершенно отличны от миелина. Состав фосфолипидов обогащенных фракций нейронов и нейроглии коры мозга крысы представлен в табл. 4.3.
Чем более анатомически дифференцированно подходить к нервной ткани, тем больше различий обнаруживается в липидном составе, поскольку функционально различные нейрональные и глиальные клетки имеют своеобразный липидный состав. В состав большинства липидов входят жирные кислоты.
В мозге они гораздо разнообразнее, чем в других тканях. Это намного увеличивает число индивидуальных липидов мозга. Содержание жирных кислот в головном мозге гораздо выше, чем в других органах, и составляет примерно 20-25% в расчете на сухую массу ткани. Разнообразие жирных кислот в этом органе поразительно. Применение газожидкостной хроматографии позволило продемонстрировать наличие в головном мозге более 50 жирных кислот с длиной цепи от 12 до 26 углеродных атомов, среди которых найдены насыщенные, ненасыщенные, нормальные, гидроксизамещенные, нечетные и др. Ненасыщенные кислоты мозга могут содержать от 1 до 6 двойных связей. Особенностью, липидов мозга является относительно большое содержание длинноцепочечных полиеновых кислот 20:4, 22: 5, 22:6. Отдельные классы и фракции липидов мозга характеризуются своим набором жирных кислот. Имеет место также определенная специфичность жирнокислотного состава в липидах разных отделов мозга, разных типов его клеток, субклеточных структур.
Таблица 4.2. Состав липидов основных типов нервных клеток мозга крысы (мкмоль/мг сухой массы)
|
Липиды |
Нейроны |
Астроглия |
Олигоден-дроглия |
Миелин |
|
Холестерин |
6,610 |
14,100 |
10,800 |
54,900 |
|
Цереброзиды |
0,513 |
0,689 |
2,610 |
22,000 |
|
Сульфатиды |
0,090 |
0,142 |
0,472 |
2,890 |
|
Общие фосфолипиды |
22,400 |
35,600 |
23,400 |
41,800 |
|
Ганглиозиды |
0,223 |
0,582 |
0,239 |
0,0453 |
|
Молярное отношение: холестерин – цереброзиды-фосфолипиды |
1:0,075:3,5 |
1:0,05:2,5 |
1:0,25:2,2 |
1:0,40:0,76 |
Иллюстрацией этого могут служить данные табл.4.4, где приведен жирнокислотный состав фосфолипидов синаптосом и миелина — двух разных типов мембранных структур ЦНС резко различающихся по своему происхождению и функциям.
Основные фосфотриглицериды мозга и точки гидролиза фосфолипазами показаны на рисунке
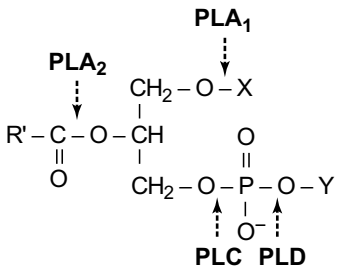
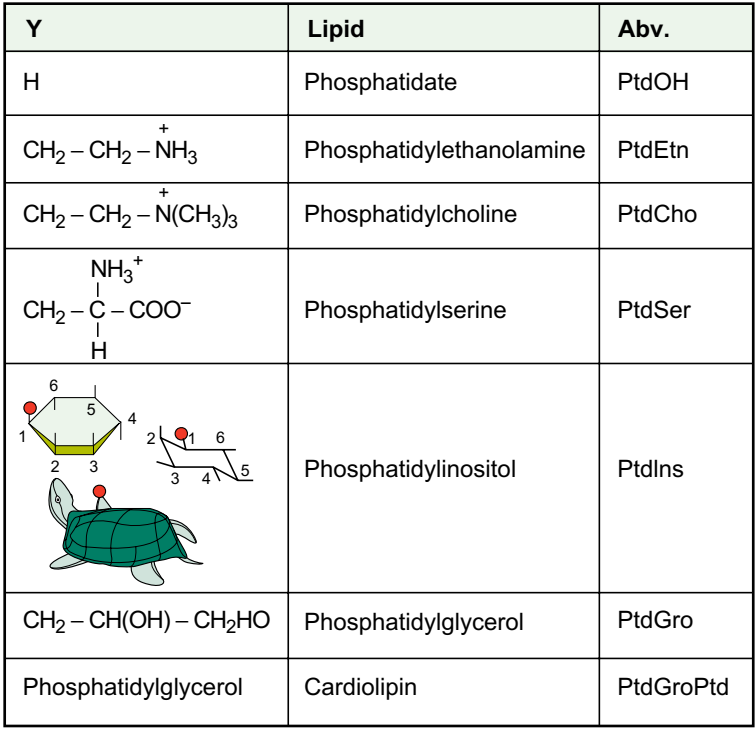
На рисунке структура фосфоглицеридов. В большинстве липидов Х представляет собой ацил, то есть R (C). В алкиловых эфирах, присутствующих главным образом в мозге этаноламинфосфоглицериды (2 - 3%), X является длинноцепочечным углеводородом (C16, C18). Для плазмогагенов, составляющих около 60% взрослого человеческого мозга PtdEtn, X представляет собой 1-алк-1-енил (то есть CH CH- R). Стрелки указывают места ферментативного гидролиза фосфоглицеридов. PLA1, фосфолипаза A1; PLA2, фосфолипаза A2; PLC, фосфолипаза C; PLD, фосфолипаза D. Обратите внимание, что мио-инозитол записывается в d-конфигурации, где 1-я позиция связана с частью PtdOH. Для полифосфоинозитидов в 3, 4 или 5 положениях присутствуют дополнительные фосфатные группы. Стереохимии инозита принято рассматривать с использованием представлений черепахи.
Таблица 4.3. Содержание индивидуальных фосфолипидов в коре мозга крысы
|
Фосфолипиды |
Нейроны |
Нейроглия |
|
Лизофосфатидилхолин |
3,9 |
1,9 |
|
Фосфатидилхолин |
46,1 |
46,9 |
|
Сфингомиелин |
6,7 |
9,5 |
|
Фосфатидилсерин |
9,1 |
7,1 |
|
Фосфатидилинозит |
7,7 |
5,9 |
|
Фосфатидилэтаноламин |
25,1 |
24,9 |
|
Фосфатидная кислота |
1,8 |
3,6 |
В синаптосомах велико содержание жирных кислот — С 22:6, а в миелине высок процент моноеновых кислот — 18:1. Возможно, что высокое содержание докозагексаеновой кислоты в синаптосомах необходимо для активного транспорта ионов, так как активность №+, К+-АТФазы в них зависит от присутствия полиеновых кислот в фосфолипидах. В мозге имеются регуляторные механизмы, поддерживающие степень ненасыщенности и специфичность жирнокислотного состава в липидах.
Таблица 4.4. Состав жирных кислот фосфолипидов синаптосомальных и миелиновых мембран коры мозга обезьяны (% от общего содержания кислот)
|
Шифр жирной кислоты |
Фосфатидилхолин |
этаноламин |
серин+монофосфоинозитид |
|||
|
|
синаптосомы |
миелин |
синаптосомы |
миелин |
синаптосомы |
миелин |
|
18:0 |
12,4 |
17 |
25,5 |
15,9 |
44,3 |
43 |
|
18:1 |
27,2 |
42,3 |
12,1 |
33,2 |
11,4 |
38,5 |
|
20:1 |
0,7 |
0,9 |
1,6 |
' |
9,3 |
2,5 |
|
20:4 |
3,8 |
3,2 |
10,1 |
11,6 |
8,3 |
6,3 |
Изменение жирнокислотного состава приводит к нарушению функциональной деятельности мозга.
Таблица 4.5. Распределение жирных кислот в фрагментах диацилглицерина фосфоглицеридов мозга крыс
|
C-1 |
C-2 |
PtdIns (mol %) |
PtdIns4,5P2 (mol %) |
PtdCho (mol %) |
PtdEtn (mol %) |
PtdSer (mol %) |
|
16:0 |
22:6 |
1.4 |
0.1 |
3.3 |
4.8 |
0.8 |
|
16:0 |
20:4 |
7.8 |
9.5 |
4.4 |
2.3 |
0.6 |
|
18:1 |
20:3 |
4.1 |
1.1 |
Tr |
Tr |
Tr |
|
18:0 |
22:6 |
Tr |
1.0 |
2.5 |
17.6 |
42.4 |
|
14:0 |
16:0 |
0.6 |
0.4 |
3.1 |
1.5 |
0.8 |
|
18:0 |
22:5 |
1.0 |
0.7 |
0.4 |
0.2 |
5.3 |
|
18:0 |
20:4 |
49.5 |
66.1 |
3.8 |
22.5 |
3.8 |
|
18:1 |
18:1 |
1.7 |
2.1 |
3.4 |
11.1 |
7.0 |
|
16:0 |
18:1 |
12.7 |
6.5 |
36.2 |
15.8 |
9.1 |
|
16:0 |
16:0 |
6.9 |
1.4 |
19.2 |
0.7 |
Tr |
|
18:0 |
18:1 |
7.0 |
4.6 |
14.1 |
14.8 |
23.7 |
PtdIns, фосфотидилинозитол; PtdIns(4,5)P2, фосфотидилинозитол-4,5-дифосфат; PtdCho, фосфатидилхолин; PtdEtn, фосфатидилетаноламин; PtdSer, фосфатидилсерин.
Строение основных сфинголипидов мозга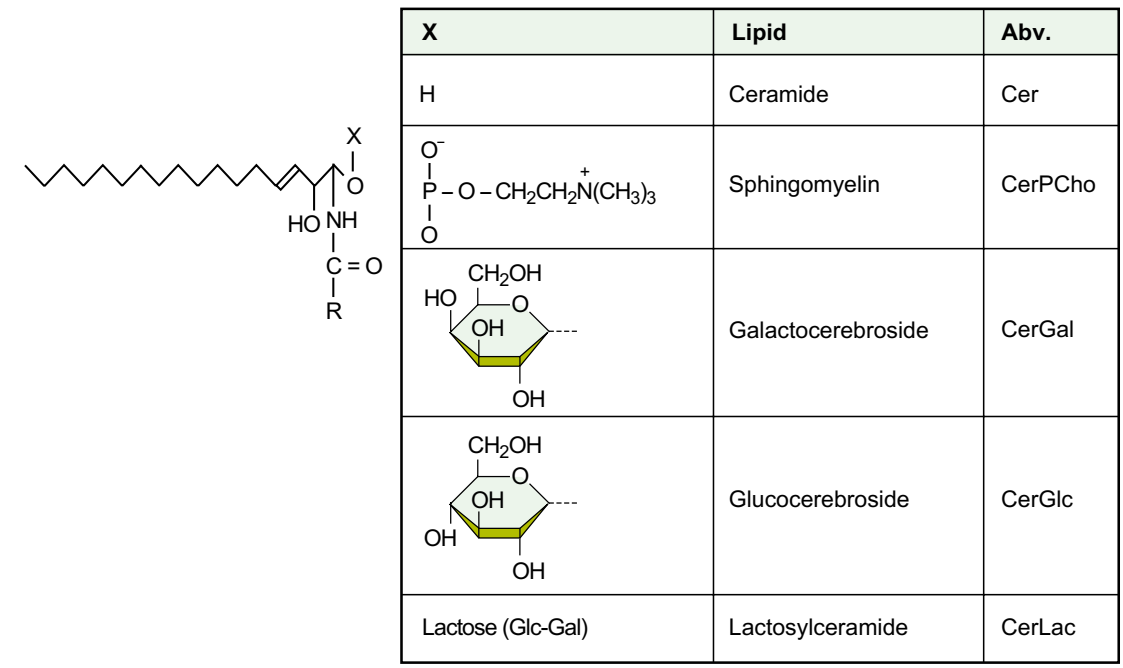
Рис.Структура некоторых простых сфинголипидов. X может быть сложным полисахаридом, содержащим сиаловую кислоту (ганглиозиды) или нет (глобозиды). См. Также следующие рисунки 3-4 и 3-9 для понимания номенклатуры и структуры некоторых сложных сфинголипидов мозга.
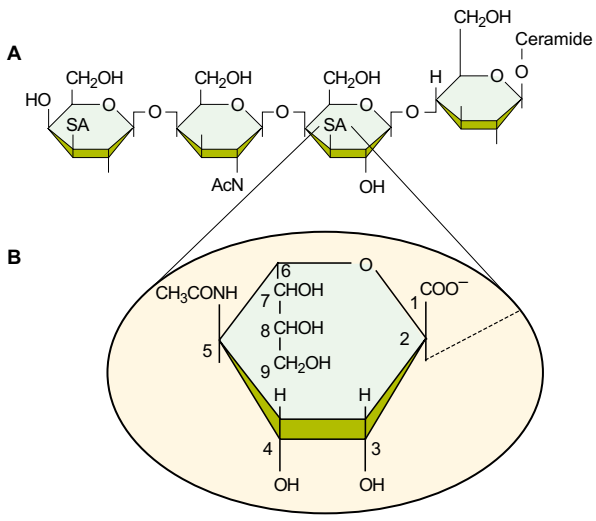
(A) Структура основного ганглиозида головного мозга, который называется GD1a в соответствии с номенклатурой Svennerholm. G обозначает ганглиозид, D обозначает дисдиало, 1 относится к основному основанию тетрасахарида (Gal-GalNac-GalGlc-) и отличает позиционные изомеры в терминах расположения остатков сиаловой кислоты (см. Также фиг.3-9). В номенклатуре IUPAC-IUB этот ганглиозид называется IV3NeuAc, II3NeuAc-Gg4Cer, где римские цифры обозначают сахарный фрагмент (из церамида), к которому прикреплены сиаловые кислоты (NeuAC), а аравийский цифровой надстрочный символ обозначает положение в сахарном фрагменте где NeuAC прилагаются; Gg относится к сериям ганглио (Gal-GalNAc-Gal-Glc) и индексу 4 к четырехуглеводной магистрали для серии «ганглио». (B) Структура сиаловой кислоты, также называемая N-ацетилнеураминовой кислотой (NeuAc или NANA). Ганглиозиды мозга человека - все производные N-ацетила; однако некоторые другие млекопитающие, такие как коровы, в мозгу могут содержать N-гликолильные производные. Предшественником метаболического биосинтеза для сиалилирования гликоконъюгатов является CMP-сиаловая кислота, образующая фосфодиэфир 5'OH цитидина и 2-положение нейрамининовой кислоты.