Введение в нейробиологию
12. Мембранный транспорт и генерация мембранных потенциалов
12.4. V0V1-ПРОТОННЫЕ НАСОСЫ
V-ATPase накачивает протоны в органеллы, из Голджи. Его специализированная нейронная функция - генерировать электрохимический градиент, который активирует H + -антипортеры, которые загружают нейротрансмиттеры в пресинаптические везикулы (глава 10). Структура V0V1 этих насосов (фиг.5-9) аналогична структуре синтаз F0F1-ATP и они прокачивают протоны с помощью аналогичного ротора и механизма статора. В отличие от F0F1, животные которого выражаются только в митохондриях, особая органелла Гольджи, цели которой V0V1 зависит от изоформы субъединицы a, которая экспрессируется [33].
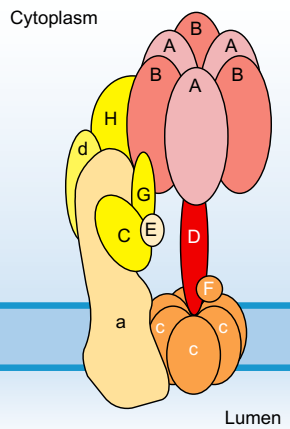
РИСУНОК 5-9 Структурная организация везикулярного протонного насоса V-ATPase: V-ATPase состоит из множества субъединиц, организованных, как показано цветовым кодированием: цитоплазматический домен V1 содержит восемь различных субъединиц (A-H) с тремя копиями связывающих субъединиц А и В АТФ; мембранный домен, называемый V0, представляет собой гексамерное кольцо, состоящее из субъединиц протеолипидов (5 c и 1 c '') и одиночных копий субъединиц a и d. Функционально V-ATPase состоит из ротора (белая надпись) и домена статора (черная надпись). Ротор состоит из гексамерного протеолипидного c-кольца и стебля из субъединиц D и F; статор состоит из оставшихся субъединиц, которые прикреплены к мембране через субъединицу a. Экспрессия различных субъединиц изоформы преследует VATPase для разных мембран. АТФ-связывание и гидролиз тремя субъединицами A, по-видимому, действуют на субъединицу D, чтобы индуцировать вращение через асимметричные конформационные переходы, как описано для синтазы F0F1 ATP. Вращение гексамерного c-кольца транслоцирует протоны из цитоплазматической границы гексамера протеолипида в просвет синаптических или везикул, полученных из Гольджи. Область V1 обратимо диссоциирует в физиологических условиях для регулирования активности V-АТФазы. (С разрешения ссылки [86].)