Дисциплина Экспериментальная патобиохимия клетки
Медиаторы воспаления
5. Пироптоз
5.2. Молекулярные механизмы
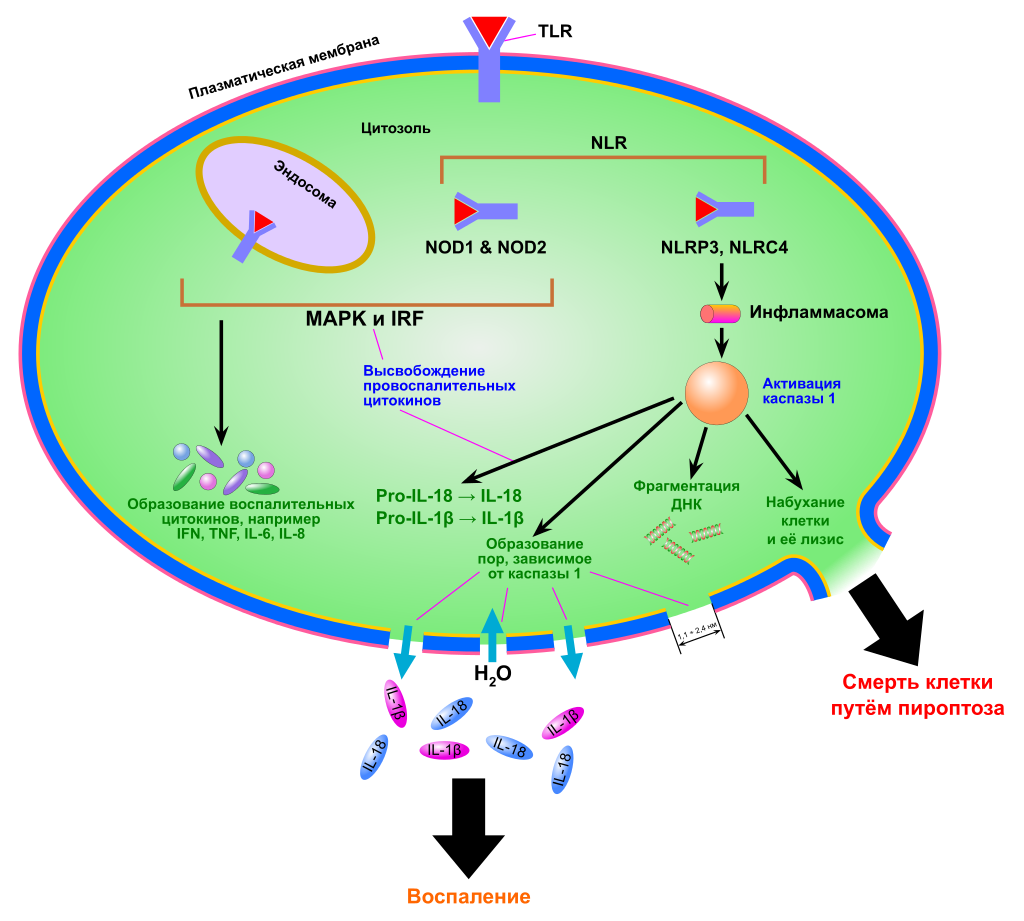
Пироптоз запускается сигналами опасности, которые распознаются во вне- и внутриклеточной среде двумя группами рецепторов опознавания паттерна: Nod-подобными рецепторами (NLR), находящимися в цитоплазме, и Toll-подобными рецепторами (TLR), расположенными в плазматической мембране. Эти сигналы опасности могут выделяться патогенными организмами, при повреждениях тканей. При связывании с NLR внутриклеточных бактериальных, вирусных или принадлежащих самой клетке сигналов опасности начинается сборка мультибелкового комплекса — инфламмасомы (от англ. inflammation — воспаление). Сборка инфламмасомы приводит к активации каспазы 1, которая необходима для образования и выделения провоспалительных цитокинов. Наиболее хорошо изученная инфламмасома, NLRP3, имеет домены трёх главных типов: домены, содержащие обогащённые лейцином повторы (LRR), центральный нуклеотид-связывающий домен олигомеризации (NBD) и N-концевой пириновый домен (PYD). Взаимодействие каспазы 1 и NLRP3 осуществляется при помощи адаптерного белка ASC. ASC содержит домен активации и рекрутирования каспаз (CARD-домен), который связывается с прокаспазой 1 и облегчает её активацию через взаимодействие с её доменом CARD[9]. В результате прокаспазы 1 сближаются, димеризуются и разрезают друг друга на фрагменты p10 и p20, которые способы осуществлять процессинг про-IL-1β и про-IL-18. В других случаях инфламмасома активирует каспазу 1 через другие белки, содержащие домен CARD или PYD[10].
Ключевая роль каспазы 1 в пироптозе была продемонстрирована в 1995 году в экспериментах с мышами[en]*, нокаутными по каспазе 1: их клетки были неспособны к пироптозу. Впрочем, в 2011 году было показано, что все эти мыши несли ещё и сопутствующую мутацию в гене каспазы 11[en]. Поэтому каспаза 11 (человеческие ортологи — каспазы 4 и 5 также играет важную роль в пироптозе и врождённой защите от внутриклеточных патогенов. Более того, каспаза 11 может усиливать активацию каспазы 1 при инфекциях, вызванных грамотрицательными бактериями, так что обе эти каспазы могут инициировать пироптоз.
Для прохождения пироптоза необходима протеолитическая активность каспаз, и при пироптозе происходит разрушение или нарушение работы ряда белков, необходимых для функционирования и выживания клетки. К настоящему моменту известно множество мишеней каспазы 1. Среди них белки цитоскелета, внутриклеточного транспорта, трансляции и ключевых метаболических путей (например, гликолитические ферменты альдолаза, триозофосфатизомераза[en], глицеральдегид-3-фосфатдегидрогеназа[en], енолаза и пируваткиназа). Кроме того, каспаза 1 процессирует и другие каспазы: 4, 5 и 7. Хотя каспаза 7 является эффекторной каспазой апоптоза, её активация для пироптоза необязательна, что свидетельствует о различиях в сигнальных путях этих двух видов клеточной гибели.
Ранним этапом пироптоза является образование небольших пор в клеточной мембране, проницаемых для катионов. Это нарушает ионный баланс клетки и приводит к её набуханию и лизису. Через эти поры в клетку входят ионы Ca2+, которые участвуют во многих клеточных событиях, связанных с каспазой 1. Лизису клетки предшествуют конденсация ядра, фрагментация ДНК[en], а также секреция IL-1β и IL-18. Кальций также способствует экзоцитозу лизосом, которые «закрывают» прорехи в мембране и делают возможным высвобождение антимикробных соединений. Эти соединения убивают бактерий, находящихся во внутриклеточной среде. Кроме того, путём экзоцитоза до лизиса клетки наружу выделяются фагоцитированные частицы и внутриклеточные патогены.
В ходе пироптоза из клетки высвобождаются так называемые молекулярные паттерны, связанные с повреждениями (англ. Damage-associated molecular patterns, DAMP), которые во внеклеточной среде выступают сильными провоспалительными факторами. К их числу принадлежат АТФ, ДНК, РНК, белки теплового шока. DAMP запускают воспалительные процессы в клетках, активируют выделение цитокинов и усиливают нацеливание Т-клеток на определённые мишени. Один из DAMP, HMGB1, играет особую роль в пироптозе. HMGB1 — это ядерный транскрипционный фактор, который высвобождается наружу при пироптозе и активирует клеточные рецепторы TLR4 и RAGE, которые, в свою очередь, активируют выделение цитокинов и миграцию клеток. Показано, что один только HMGB1, без IL-1β, IL-18 и других DAMP, может вызывать воспаление.
Сигнальные липиды группы эйкозаиноидов, такие как лейкотриены и простагландины, увеличивают проницаемость кровеносных сосудов и способствуют воспалению. В 2012 было показано, что входящий ток кальция после активации каспазы 1 может индуцировать синтез эйкозаиноидов. При этом глицин, ингибирующий лизис клетки, но не секреторные пути, связанные с каспазой 1, не блокировал выделение эйкозаноидов, подтверждая, что они действительно секретируются, а не просачиваются во внешнюю среду пассивно при увеличении проницаемости мембраны. Сигнальные пути эйкозаиноидов не всегда связаны с активацией каспазы 1, однако они представляют собой очень быстрый и сильный провоспалительный сигнал, запускаемый каспазой 1.
В ходе ряда исследований было показано, что активация каспазы 1 и аутофагия антагонистичны по отношению друг к другу: активная каспаза 1 подавляет аутофагию, а аутофагия мешает активации каспазы 1 и уменьшает количество IL-1β и инфламмасом в цитозоле.